Persistance des “races humaines” en Sciences de la Vie – Partie 2
Les variations graduelles de l’information génétique des individus avec la distance géographique ont démonté l’idée que l’on pouvait classer génétiquement les individus en groupes nommés “races humaines” (voir précédent article). Mais dans le domaine scientifique, en jouant sur les termes employés, notamment avec la polysémie du terme de “race”, et sur certaines représentations graphiques, certains chercheurs permettent à cette idéologie des “races humaines” de se maintenir.
Races ou sous-espèces ?
On observe que le terme de “race” est utilisé en biologie pour regrouper des individus au sein de certaines espèces animales domestiquées (“races domestiques” de chiens, de bovins…) ou parfois pour regrouper des individus au sein d’espèces naturelles (expression de “race géographique”) bien que dans ce cadre naturel, c’est le terme de “sous-espèce” qui est généralement utilisé.
Dans le cadre des espèces domestiques, les races sont des groupes formés artificiellement : l’Homme impose une barrière génétique en empêchant deux individus d’une même espèce de se reproduire entre eux (de se “croiser”), selon certains critères. Ces critères peuvent être “esthétiques”, comme un chat avec un pelage particulier (telle la race Bengal avec des tâches de type “léopard”), ou reposer sur des besoins en terme de production alimentaire (comme la race de vaches Prim’Holstein spécialisée dans la production de lait ou la race Charolaise spécialisée dans la production de viande).
Dans le cadre des espèces naturelles, l’Homme, pour essayer de comprendre comment de nouvelles espèces se forment naturellement, utilise le terme de “sous-espèce” : deux groupes d’une même espèce peuvent évoluer en deux sous-espèces puis en deux espèces. Ainsi les girafes Masai sont interfécondes et peuvent se reproduire naturellement entre elles (il s’agit d’une même espèce), mais dans le nord de la Tanzanie, un rift sépare géographiquement des groupes de girafes Masai formant deux sous-espèces (les girafes sont interfécondes mais elles ne peuvent se reproduire dans la nature : une barrière génétique existe, liée à la géographie).
Au vu de ces usages, on justifie parfois l’inexistence des “races humaines” en expliquant que l’on ne peut faire des groupes nommés “races” seulement au sein des espèces domestiques. Mais la tentation de définir des “races humaines” peut alors se camoufler derrière des regroupements en “sous-espèces humaines”. De fait, rappelons que d’un point de vue biologique, seuls les individus existent et que sur cette base tout regroupement d’individus (à l’exemple de la notion d’espèce) est un concept utile (voir l’article précédent “Persistance des « races humaines » en sciences de la Vie, Partie 1” dans la revue précédente) et l’on pourrait décliner le terme de “races” en races domestiques et races naturelles ou le terme de “sous-espèces” en sous-espèces domestiques et sous-espèces naturelles.
Pour combattre l’idée d’existence de “races humaines”, le plus efficace et clair en terme de classification est de considérer, comme nombre de chercheurs le font, que les termes races, sous-espèces et variétés recouvrent la même signification, avec un usage du terme race propre aux animaux domestiques et du terme variété propre aux plantes domestiques (avec leurs spécificités).
Ainsi en biologie, deux groupes forment deux sous-espèces (ou deux races) au sein d’une espèce s’ils présentent une différenciation génétique relativement importante entre eux et qu’il y a des barrières génétiques nettes entre eux (donc pas de variations génétiques lentes et graduelles).
Degré de différenciation entre deux groupes
La catégorie de sous-espèce ou de race est utile si elle obéit à des critères mesurables. Le degré de différenciation génétique entre deux groupes est ainsi mesuré à partir des petites différences génétiques entre individus (soit à partir des 0,1 % d’ADN environ qui différencient deux individus humains). Mais dans la vulgarisation scientifique, le degré de différenciation entre deux groupes est souvent donné comme seul argument pour soutenir soit l’absence soit l’existence de “races humaines”, ce qui entraîne des batailles de chiffres qui ne font que semer le doute (1). Or les deux conditions, de différenciation et de frontières génétiques nettes, sont fondamentales.
La méthode traditionnelle considère que deux groupes sont deux sous-espèces s’ils présentent un degré de différenciation génétique empirique globalement supérieur à 25 % et que la frontière génétique est nette entre les deux groupes. Par exemple au sein de l’espèce humaine, on trouve une augmentation continue relativement lente de l’indice de différenciation entre deux populations quand la distance géographique entre ces deux populations augmente : une augmentation entre 0 et plus de 20 % sur près de 30 000km (2). Il n’y a donc pas de sous-espèces dans l’espèce humaine (en lien avec le processus principal de différenciation génétique entre populations, l’isolement par distance). Par comparaison les populations de deux sous-espèces de girafe Masai sont présentes dans le nord de la Tanzanie, de part et d’autre de la falaise du rift Est-Africain, avec un indice de différenciation soit faible (0,5 %) soit fort (de 19 à 37 %) (3).
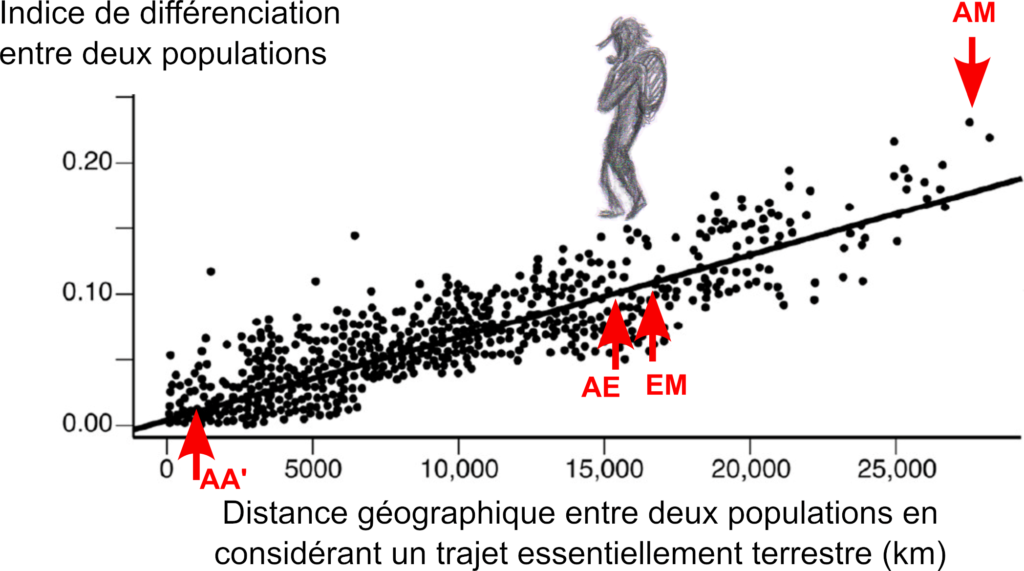
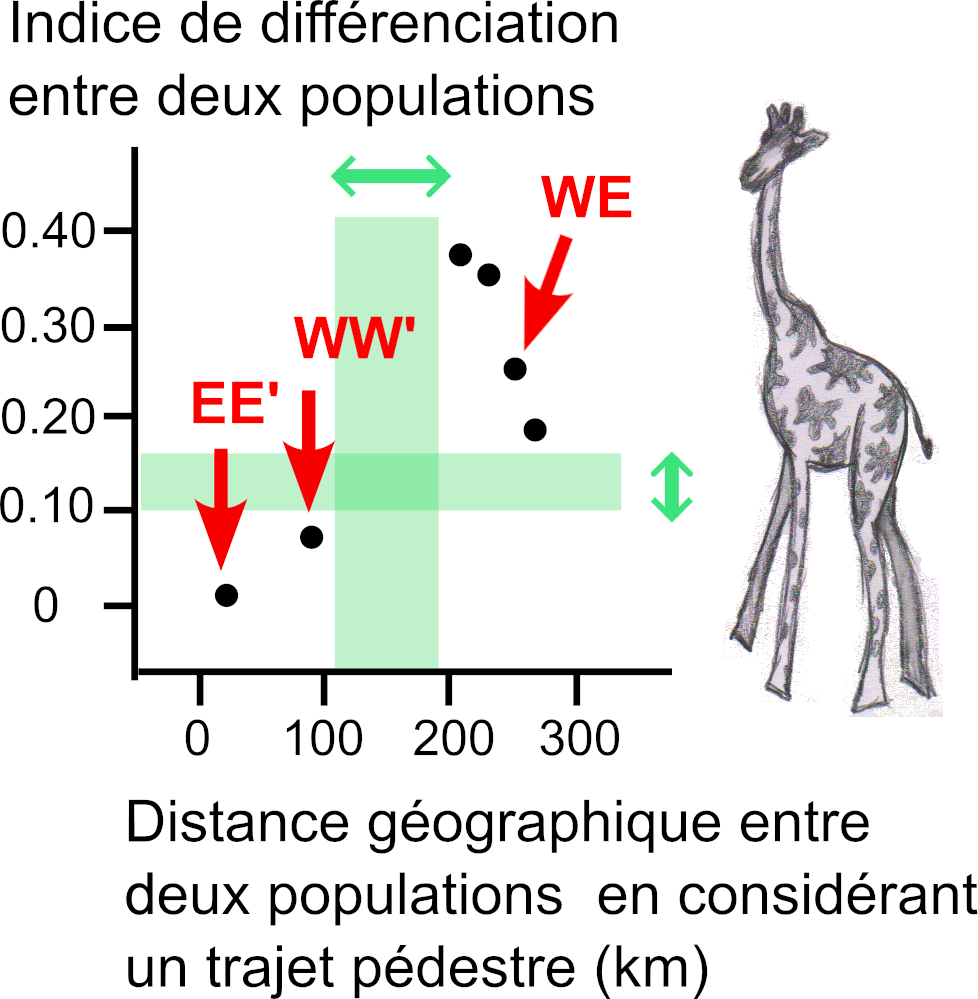
Évolution de l’indice de différenciation en fonction de la distance géographique entre deux populations. L’“indice de différenciation” entre deux populations (ou “indice de fixation”, créé dans les années 70) est un outil qui se fonde sur les fréquences des variants (allèles) de séquences d’ADN de ces deux populations.
Gauche : cas de l’espèce humaine. Exemple de lecture : la population A localisée en Afrique a un indice de différenciation proche de zéro avec une population A’ localisée également en Afrique (point AA’), un indice de 10 % avec une population E localisée en Europe (point AE), un indice de plus de 20 % avec une population M localisée en Malaisie (point AM). Cette population M, a un indice de 10 % la population E localisée en Europe (point EM). D’après Ramachandran et al. 2005 (données HGDP-CEPH, séquences d’ADN de type microsatellites – tous les indices ne sont pas représentés) (2). En moyenne 15 % de la variabilité génétique se trouve entre deux populations et 85 % au sein d’une même population.
Droite : cas de quatre populations de l’espèce de Girafe Masai localisées dans le nord de la Tanzanie. Les populations E et E’ sont localisées à l’est du rift et ont un faible indice de différenciation (point EE’), de même pour les populations W et W’ localisées à l’ouest du rift (point WW’) ; par contre cet indice est fort entre les autres couples de populations, comme W et E (point WE). Les doubles flèches claires indiquent la barrière génétique (flèche verticale) causée par la barrière géographique (flèche horizontale). D’après les données de Lahay et al. (3).
Le seuil de 25 %, empirique, est discutable car arbitraire, mais le critère indissociable de frontière génétique nette permet de lever ce problème. En outre, le développement du séquençage génétique et des outils mathématiques et numériques en biologie a permis le développement de nouvelles méthodes (comme celle fondée sur la lignée évolutive) : aujourd’hui c’est le croisement de ces différentes méthodes qui permet de déterminer l’existence de sous-espèces au sein d’une espèce. Des méthodes qui ont pu bouleverser des classifications (comme le passage d’une espèce de girafes à quatre espèces de girafes), mais rien de tel au sein de l’espèce humaine !
Un arbre pour représenter l’histoire humaine ?
Graphiquement, il y a plusieurs façons de représenter le degré de différenciation génétique entre deux populations échantillonnées. On peut simplement représenter des lignes dont la longueur dépend du degré de différenciation entre les deux populations. On peut également interpréter ces degrés de différenciations génétiques en terme de temps depuis lequel deux groupes ont divergé à partir d’un ancêtre commun, et les représenter sous la forme d’un arbre. Au sein d’une espèce, une divergence implique alors la formation de deux groupes distincts, et donc de deux sous-espèces (ou races). Chaque groupe correspond à une lignée évolutive.
Au sein de l’espèce humaine, de tels arbres sont donc faux scientifiquement (tester des arbres ou la robustesse des lignées évolutives constitue une méthode incontournable pour déterminer l’existence ou non de races ou sous-espèces). Ceci est expliqué par le fait que tout au long de l’histoire humaine, les mélanges ont été incessants (tout comme ces mélanges expliquent les variations graduelles de la diversité humaine).
Pourtant de tels arbres sont encore courants dans les articles scientifiques. A. Templeton, avec d’autres, à juste titre explique que “cette pratique est indéfendable. Elle est également socialement irresponsable”. Une représentation plus juste si l’on veut que la représentation graphique retrace une évolution historique serait sous forme de treillis ou de nervures de feuilles (2).
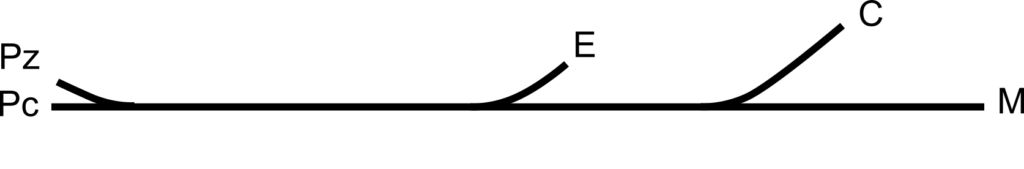
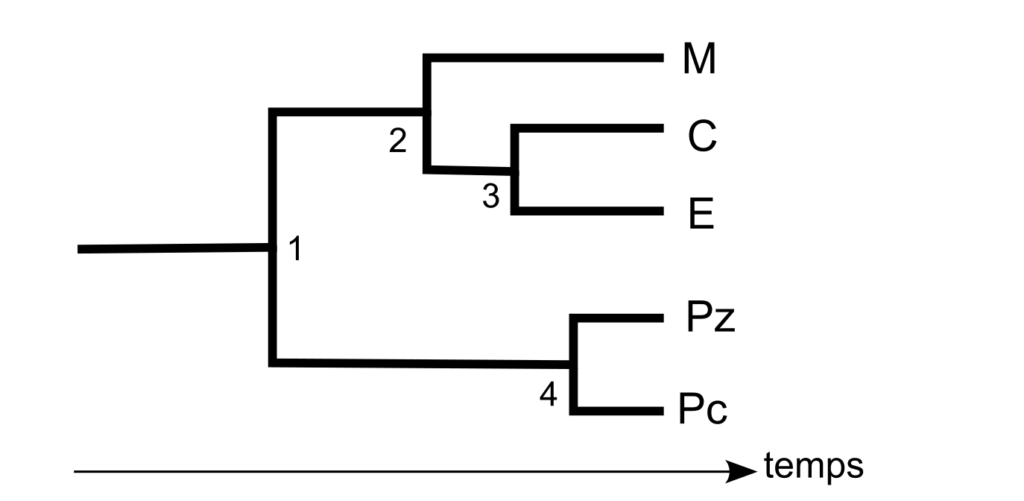
Représentation du degré de différenciation entre cinq échantillons d’individus (4). Représentation simple (en haut) et représentation en interprétant un lien d’ascendance (en bas, les ancêtres communs sont numérotés de 1 à 4), ce qui suggère une séparation qui est erronée entre les populations. Échantillons d’individus vivant : au Zaïre (Pz), en Centrafrique (Pc), en Europe (E), en Chine (C), en Malaisie (M). D’après Cavalli Sforza et al. 1996 et (2). Dans les deux cas, l’échantillonnage est discontinu et les choix effectués (comme la représentativité qu’auraient quelques individus de “l’Europe”) peuvent et doivent être critiqués.
La représentation de tels arbres (faux scientifiquement) au sein de la lignée humaine est encore présente dans certains supports de documents mis à disposition des élèves ou des enseignants de SVT dans l’Éducation nationale (5). Elle a pour cause la répercussion d’une pratique erronée dans certaines publications scientifiques et la confusion avec la méthode cladistique qui utilise de tels arbres, mais pour grouper des espèces : dans ce cas, cela est juste car par définition deux individus de deux espèces différentes ne sont pas interféconds ; il y a bien eu une division d’un groupe ancestral en deux nouvelles espèces. Ce qui n’est pas le cas de la population humaine en sous-groupes. En outre l’assimilation d’échantillons d’individus considérés comme “référence” à la nationalité d’un pays ou d’un continent entretient également les confusions.
Définitions communes et définition biologique
Au cours de l’histoire, le terme race a eu plusieurs significations et usages, avec un dénominateur commun : il s’agit de désigner un groupe d’individus, au sein d’une espèce ou correspondant à une espèce. Si l’on met de côté les caractérisations morales que le terme race véhicule parfois, le terme de lignée (d’ascendance ou descendance) est presque toujours sous-jacent au terme de race. Ces deux idées sont reprises dans la définition biologique du terme “race” et dans les définitions communes du terme “races humaines”.
Ainsi, la définition traditionnelle (du XIXe siècle) de “race humaine” repose sur des caractères visibles liés à des différences socio-culturelles, se superposant aux continents (sous-entendant des “lignées continentales”). Avec le développement de la génétique, cette définition s’est modifiée dans l’esprit de certains chercheurs et du grand public pour délimiter des groupes au sein de l’espèce humaine présentant des “différences génétiques moyennes” qui se superposeraient avec les continents ou une partie d’entre eux. Parfois des caractères visibles sont aussi associés à cette définition. En outre, des liens avec des maladies et/ou des différences sociales peuvent être évoquées.
Certes, les deux chercheurs en biologie, B. Jordan et D. Reich critiquent clairement le racisme du diplomate et essayiste du XIXe siècle Gobineau ou celui du chercheur codécouvreur de la structure de l’ADN, J. Watson. Mais parler de l’existence de “groupes apparentés génétiquement entre eux” correspondant globalement “aux catégories « raciales »” (B. Jordan (6)), ou de “différences génétiques moyennes entre « races »” (D. Reich (7)), revient à utiliser dans le domaine scientifique de la biologie une autre définition du terme “race” que celle définie… en biologie ! Ceci témoigne a minima d’un énorme manque de rigueur scientifique et d’une influence de l’idéologie racialiste sur leur travail scientifique (l’idéologie racialiste consiste à faire des races dans l’espèce humaine sans qu’il n’y ait d’idée de hiérarchie entre elles).
La notion de race en sciences de la Vie reprend ainsi l’idée de descendance au sein d’une espèce avec présence de barrières génétiques entre groupes, idée que l’on retrouve dans les différentes définitions communes du terme “race”. Or la confrontation des données biologiques dans l’espèce humaine à cette définition aboutit à infirmer l’existence de races au sein de l’espèce humaine. Face à cela, l’idéologie racialiste cherche à s’imposer en Sciences de la Vie par l’utilisation du terme “races humaines” ou de termes annexes (comme “groupes continentaux”, “groupes d’ascendances”) comme s’il s’agissait de données allant de soi. C’est notamment sur de tels courants idéologiques dans la recherche que s’appuient certaines entreprises pour vendre leurs produits, ce que développera le prochain article.
Laure Jinquot
Sources :
- (1) Ainsi dans le livre On vient tous vraiment d’Afrique ? par E. Heyer et C. Reynaud-Paligot, Flammarion, 2019, on peut lire “si l’on compare deux chiens pris au hasard dans deux races différentes, 30 % des différences sont dues aux différences entre les races. Ce chiffre est à comparer avec les 5 % des différences que l’on trouve entre deux humains qui sont dues au fait qu’ils viennent de deux populations très éloignées sur la planète” et d’en déduire donc qu’il n’y a pas de “races humaines”. Outre que les chiffres ne sont pas sourcés, il est assez facile de trouver des articles dans la littérature qui donnent de faibles indices de différenciation (inférieurs à 10 % ou 5 %) entre deux races domestiques : côté chiens, chats, ânes (voir sources ci-après)… La raison en est que la création d’une race domestique passe par l’absence de croisement imposée par l’Homme (barrière génétique) sur la base de caractères d’intérêts que ceux-ci se traduisent ou non par une différenciation génétique forte entre deux groupes (différenciation qui augmentera avec le temps). En outre, le seuil empirique de 25 % (fort degré de différenciation) a été fixé dans le cadre d’observations naturelles de sous-espèces. D’où l’importance de considérer une “différenciation relativement importante” entre deux groupes et une “barrière génétique nette”.
- Exemple de sources : côté chiens Mitochondrial DNA Sequence Variation in Portuguese Native Dog Breeds : Diversity and Phylogenetic Affinities par Pires et al., Journal of Heredity, 2006 ; côté chats Patterns of molecular genetic variation among cat breeds par Menotti-Raymond et al., janvier 2008, Science Direct ; côté ânes Mitochondrial DNA genetic diversity in six Italian donkey breeds par Cozzi et al., mars 2017, Mitochondrial DNA part A.
- (2) Templeton : The genetic and evolutionary significance of human races, 2002 et Biological Races in Humans, 2013.
- (3) Genetic evidence of population subdivision among Masai giraffes separated by Gregory, Rift Valley in Tanzania par Lohay et al., mai 2023, Ecology et Evolution.
- (4) Le degré de différenciation entre deux populations peut également être déduit de la distance génétique entre deux populations de référence (fondée sur le calcul de distance génétique entre individus). On peut prendre pour exemple de lecture un degré de différenciation de 20 % entre Pc et M et de 10 % entre PC et E ou entre Pc et M. Alors le graphique du haut est correct car il montre bien que la distance entre Pc et M est deux fois plus grande que la distance entre Pc et E ou entre E et M. Par contre, dans le graphique du bas, la distance horizontale entre M et Pc est la même que celle entre E et Pc ce qui signifie que le degré de différenciation est le même entre M et Pc qu’entre E et Pc, ce qui est faux ! Problème auquel s’ajoute l’idée fausse de division d’un groupe à partir d’un ancêtre commun (par exemple la population ancestrale 3 donnant les populations C et E).
- (5) Voir par exemple le livre scolaire d’enseignement scientifique Le livre scolaire, p 212 (doc. 3 : Des hybridations entre populations d’humains modernes néandertaliens : https://www.lelivrescolaire.fr/page/5339681 ou un article de vulgarisation sur le site de l’ENS Lyon (acces.ens-lyon.fr) “Origine des Homo sapiens” par JC Hervé, février 2019.
- (6) La génomique et la diversité humaine, B. Jordan, 2021.
- (7) How Genetics is changing our understanding of “race” by David Reich, march 23, 2018, New York Times Opinion.
